

Важнейшим показателем функции растений служит преобразование световой энергии. К настоящему времени детально исследованы механизмы фотосинтеза, свойства основных пигментов фотосинтетического комплекса (ФСК) хлорофиллов и каротиноидов, поглощающих фотоны видимой области оптического спектра [1, 2]. Сравнительно меньше уделяется внимания другим компонентам (ДК) листа, не входящих в фотосинтетическую систему, антоцианам [3], кумаринам, сапонинам и т.п., поглощающих обычно в ультрафиолетовом (УФ) диапазоне. Соотношения интенсивностей поглощения веществами ФСК и ДК могут служить характеристиками утилизации света растениями. Указанные соотношения определяются методами абсорбционной спектрофотометрии экстрактов из зеленого листа с применением в качестве критериев числовых показателей спектров поглощения (ЧПСП) [4]. Однако ЧПСП листьев растений разных жизненных форм, в зависимости от условий обитания, хотя и могут служить их признаками и количественными показателями состояния, остаются малоизученными и недостаточно полно представлены в научной литературе.
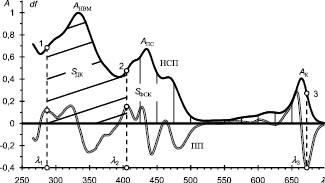
Нормированный спектр поглощения (НСП) и его первая производная (ПП) для экстракта из листьев вороньего глаза обыкновенного (Paris quadrifolia L.). 1, 2 и 3 – точки перегиба. АНВМ, АПС и АК – оптические плотности наиболее высокого, пурпурно-синего и красного максимумов. λ1, λ2 и λ3 – абсциссы точек перегиба, SФСК и SДК – площади, соответствующие поглощению веществами ФСК и другими компонентами листа. По горизонтали – длина волны λ в нм, по вертикали – оптическая плотность A и значения df производной в отн. ед. Масштаб df приведен к масштабу A для наглядности
Цель исследования
Целью работы явились сравнительные исследования и сопоставления величины ЧПСП экстрактов из листьев трав, произрастающих на открытых пространствах и под пологом леса, а также кустарников, лиан и деревьев.
Материалы и методы исследования
В работе использованы листья 25 видов деревьев, лиан, кустарников, а также трав, предпочитающих как открытые пространства («светолюбивые»), так и затененные («тенелюбивые») в фазе начала плодоношения. Для исследований брали свежие листья от 6–8 растений каждого вида во второй половине дня, из которых немедленно готовили экстракты по описанной ранее методике [4]. Спектры поглощения (СП) регистрировали на цифровом спектрофотометре UV-2501PC (Shimadzu, Япония) в диапазоне 220–710 нм с шагом 1 нм. Полученные спектры нормировали по наибольшему максимуму. Для нормированных спектров поглощения (НСП) находили первую производную и по абсциссам ее локальных экстремумов определяли длины волн точек перегиба (рисунок, точки 1, 2 и 3) контура полосы поглощения (ПП).
Для количественных оценок использовали такие ЧПСП, как коэффициент относительной фотоабсорбции (КОФ), равный отношению оптической плотности (AК) на аналитической длине волны поглощения хлорофилла в красной области (рисунок), к оптической плотности (AНВМ) на длине волны наиболее высокого максимума (НВМ):
КОФ = AК/AНВМ.
Интегральные интенсивности поглощения, численно равные площади под контуром НСП в пределах границ ПП, вычисляли по формуле определенного интегрирования Симпсона [5]. Интегральная интенсивность поглощения (SДК) компонентами листа, не входящих в состав фотосинтетического комплекса, соответствует площади под контуром ПП в УФ диапазоне (рисунок, косая штриховка). Пределами интегрирования брали абсциссу λ1 точки 1 перегиба на левом склоне контура в окрестностях AНВМ и абсциссу λ2 точки 2 перегиба на левом склоне в окрестностях «пурпурно-синего» максимума AПС.
Интегральная интенсивность SФСК поглощения веществами группы ФСК численно равна площади под контуром полосы поглощения в видимом диапазоне (рисунок, вертикальная штриховка). Для ее вычисления брали пределами интегрирования абсциссу λ2 точки 2 перегиба на левом склоне «пурпурно-синего» максимума AПС и абсциссу λ3 точки 3 перегиба на правом склоне «красного» максимума AК. Интегральные интенсивности поглощения выражали в условных единицах (у. е.).
Коэффициент относительной интегральной интенсивности поглощения (КОИП) вычисляли как отношение указанных выше интегральных интенсивностей:
КОИП = SФСК/SДК.
Нормировку спектров и все вычислительные операции производили c помощью оригинальных авторских компьютерных программ, зарегистрированных в Государственном реестре интеллектуальной собственности № 2009614442 и 2017614738 [6, 7], на обработку одного НСП затрачивали около 1-й секунды. Полученные результаты статистически обрабатывали методами малой выборки и парной линейной корреляции [8].
Результаты исследования и их обсуждение
Полученные данные (таблица) показывают, что в среднем наименьшие значения ЧПСП имеют экстракты из листьев исследованных деревьев. Среди них ель гималайская и облепиха крушиновидная, произрастающие на открытых пространствах, отличаются самыми низкими значениями ЧПСП; так, величина КОФ на 38,9–55,5, SФСК на 37,7–56,2 и КОИП на 25,8–38,7 %, а λНВМ на 2–8 нм меньше средних.
Средние значения ЧПСП экстрактов из листьев исследованных лиан мало отличаются от аналогичных показателей деревьев (p > 0,05). Среди лиан актинидия аргута и лимонник китайский, произрастающие на более светлых участках по сравнению с другими лианами, имеют наиболее низкие значения; КОФ на 15,8–26,3, SФСК на 11,8–23,6, КОИП на 25,1 %, а λНВМ на 1–3 нм меньше средних. ЧПСП остальных лиан близки к средним значениям.
Экстракты из листьев «светолюбивых» трав имеют значения ЧСПС в среднем выше, чем у деревьев, а именно: КОФ на 21,7, SФСК на 23,2 и КОИП на 11,4 %. При этом у амаранта колосистого и вероники длиннолистной, собранных на освещенных участках самые низкие значения ЧПСП на 14,7–52,9 % меньше средних, а λНВМ на 8–14 нм больше среднего. Превышение средних величин ЧПСП экстрактов из листьев кустарников над соответствующими значениями, характерными для деревьев, еще больше и достигает 37,9–45,6 %. В абсорбционных спектрах экстрактов из листьев указанных растений λНВМ находится в пределах УФ диапазона.
Наконец, самые высокие значения ЧПСП обнаружены для экстрактов из листьев «тенелюбивых» трав, у которых они превосходят аналогичные показатели деревьев в 3,13–3,48 раза, особенно у гулявника лекарственного и чемерицы даурской, произрастающих под пологом леса, при этом λНВМ находится в видимом пурпурно-синем диапазоне.
При определении взаимосвязи между ЧПСП и λНВМ получены довольно высокие значения коэффициентов корреляции от 0,95 до 0,97 (p < 0,05). Другими словами, низким значениям ЧПСП соответствуют короткие длины волн НВМ, т.е. в листьях растений с такими показателями поглощают в основном ДК в УФ диапазоне. Напротив, высоким значениям ЧПСП соответствуют поглощения в длинноволновом, видимом диапазоне, преимущественно веществами ФСК.
В спектрофотометрическом анализе экстрактов из листьев растений разных жизненных форм примененные ЧПСП ранее не использовались, поэтому возможны параллели только с известными биохимическими и физиологическими данными для фотосинтетической системы. Например, в работах, выполненных на альпийских травах [9] и саженцах тиса [10], показано, что в условиях повышенной освещенности замедляется как накопление биомассы, так и развитие растений за счет, по мнению авторов, снижения фотосинтетической активности. Исследования мезоструктуры листьев, содержания каротиноидов и хлорофиллов [9–12] свидетельствуют о возрастании потребления лучистой энергии и скорости фотосинтеза в листьях при низкой освещенности. Кроме того, в условиях высокой освещенности в листьях увеличивается содержание каротиноидов, как реакция защиты фотосистемы от фотохимического повреждения, а содержание хлорофиллов снижается [12]. Из всего этого, вместе взятого, следует, что в условиях повышенной освещенности активности фотосинтеза уменьшается, а повышение освещенности влечет за собой его возрастание. С этими следствиями вполне согласуются полученные нами низкие в 3–5 раз значения КОФ, SФСК и КОИП для растений, предпочитающих открытые пространства с повышенной освещенностью, по сравнению с растениями, произрастающими в затененных условиях.
Таким образом, можно отметить, что использованные в работе ЧПСП с большой вероятностью отображают свойства фоторецепторной системы растений, а полученные результаты согласуются с известными фактами.
Заключение
Представленный в работе экспериментальный материал демонстрирует новые подходы спектрофотометрической оценки поглощательных свойств листьев растений. Разработанный метод числовых показателей спектров поглощения можно назвать для краткости «метод ЧПСП».
Растения нижнего яруса наземной биоты закономерно отличаются наибольшими значениями ЧПСП и наиболее высокой интегральной интенсивностью поглощения. Видам растений среднего яруса присущи средние значения ЧПСП и соответственно меньшая интенсивность поглощения компонентами листа. Большинство видов растений верхнего яруса леса имеют наименьшие значениями ЧПСП и наименьшую интенсивность поглощения света.
Полученные результаты не противоречат современным представлениям биохимии и физиологии фоторецепторного аппарата растений.
Числовые показатели длины волны наибольшего максимума (λНВМ), коэффициента относительной фотоабсорбции (КОФ), интегральной интенсивности поглощения фотосинтетического комплекса (SФСК) и коэффициента относительной интегральной интенсивности поглощения (КОИП) экстрактов из листьев растений разных жизненных форм
|
Жизненные формы |
Наименование растения |
Числовые показатели спектров поглощения |
|||
|
λНВМ |
КОФ |
SФСК |
КОИП |
||
|
Деревья |
Бархат амурский Phellodendron amurense Rupr. |
292 ± 0,2 |
0,17 ± 0,02 |
27,45 ± 1,72 |
0,32 ± 0,02 |
|
Береза плосколистная Betula platyphylla Sukacz. |
266 ± 0,2 |
0,27 ± 0,02 |
37,98 ± 2,65 |
0,39 ± 0,02 |
|
|
Груша обыкновенная, сорт «Петровская» Pyrus communis L. |
284 ± 0,3 |
0,20 ± 0,02 |
28,67 ± 1,98 |
0,26 ± 0,01 |
|
|
Ель гималайская Piceae morinda Link. |
276 ± 0,2 |
0,08 ± 0,005 |
11,19 ± 0,97 |
0,19 ± 0,008 |
|
|
Облепиха крушиновидная (женская) Hippophae rhamnoides L. |
270 ± 0,2 |
0,11 ± 0,007 |
15,92 ± 1,24 |
0,23 ± 0,02 |
|
|
Черемуха Маака Padus maackii Rupr. Kom. |
282 ± 0,3 |
0,23 ± 0,01 |
32,12 ± 2,16 |
0,45 ± 0,03 |
|
|
M ± m |
278,3 ± 3,9 |
0,18 ± 0,03 |
25,55 ± 4,12 |
0,31 ± 0,04 |
|
|
Лианы |
Актинидия аргута Actinidia arguta Planch ex Miq. |
281 ± 0,3 |
0,19 ± 0,02 |
27,13 ± 1,69 |
0,32 ± 0,02 |
|
Актинидия коломикта Actinidia kolomikta Maxim. |
270 ± 0,2 |
0,14 ± 0,007 |
21,02 ± 1,65 |
0,24 ± 0,01 |
|
|
Краснопузырник округлый Celastrus orbiculata Thunb. |
268 ± 0,1 |
0,27 ± 0,02 |
37,6 ± 2,14 |
0,48 ± 0,03 |
|
|
Лимонник китайский Schisandra chinensis (Turcz.) Ball. |
268 ± 0,1 |
0,16 ± 0,01 |
24,25 ± 2,12 |
0,24 ± 0,01 |
|
|
M ± m |
271,7 ± 3,12 |
0,19 ± 0,03 |
27,5 ± 3,59 |
0,32 ± 0,06 |
|
|
Травы «светолюбивые» |
Аконит Кузнецова Aconitum kusnezoffii Rchb. |
268 ± 0,2 |
0,28 ± 0,02 |
42,84 ± 3,12 |
0,46 ± 0,03 |
|
Амарант колосистый Amaranthus viridis L. |
258 ± 0,2 |
0,22 ± 0,02 |
28,37 ± 2,34 |
0,26 ± 0,01 |
|
|
Бархатцы отклоненные обыкновенные Tagetes patula L. |
270 ± 0,3 |
0,24 ± 0,02 |
37,16 ± 2,86 |
0,44 ± 0,03 |
|
|
Вероника длиннолистная Veronica longifolia L. |
280 ± 0,4 |
0,11 ± 0,007 |
15,67 ± 0,98 |
0,24 ± 0,01 |
|
|
Горчица белая Sinapis alba L. |
256 ± 0,3 |
0,28 ± 0,01 |
42,24 ± 3,16 |
0,37 ± 0,02 |
|
|
M ± m |
266,4 ± 4,35 |
0,23 ± 0,03 |
33,26 ± 5,09 |
0,35 ± 0,04 |
|
|
Кустарники |
Жимолость съедобная Lonicera edulis Turcz. |
338 ± 0,3 |
0,25 ± 0,01 |
39,37 ± 3,02 |
0,58 ± 0,04 |
|
Ирга овальнолистная Amelanchier ovalis Medik. |
271 ± 0,2 |
0,24 ± 0,02 |
36,05 ± 3,03 |
0,35 ± 0,02 |
|
|
Сирень обыкновенная Syringa vulgaris L. |
336 ± 0,3 |
0,29 ± 0,02 |
42,04 ± 3,14 |
0,65 ± 0,05 |
|
|
Смородина красная, сорт «Ранняя сладкая» Ribes rubrum L., |
353 ± |
0,31 ± 0,02 |
43,81 ± 3,16 |
0,69 ± 0,05 |
|
|
Малина обыкновенная, сорт «Искра» Rubus idaeus L |
257 ± 0,2 |
0,37 ± 0,03 |
54,64 ± 4,02 |
0,58 ± 0,04 |
|
|
M ± m |
311,2 ± 19,5 |
0,29 ± 0,02 |
43,18 ± 3,15 |
0,57 ± 0,06 |
|
|
Травы «тенелюбивые» |
Гулявник лекарственный Sisymbrium officinale (L.) Scop. |
434 ± 0,5 |
0,59 ± 0,04 |
83,78 ± 7,32 |
0,93 ± 0,07 |
|
Каланхоэ перистое Kalanchoe pinnata (Lam.) Pers. |
434 ± 0,3 |
0,61 ± 0,05 |
90,97 ± 8,11 |
1,3 ± 0,09 |
|
|
Калужница болотная Caltha palustris L. |
435 ± 0,4 |
0,55 ± 0,04 |
84,71 ± 7,22 |
0,82 ± 0,06 |
|
|
Крапива двудомная Urtica dioica L. |
435 ± 0,4 |
0,60 ± 0,05 |
96,9 ± 8,03 |
0,80 ± 0,06 |
|
|
Чемерица даурская Veratrum dahuricum (Turcz.) Loes. |
434 ± 0,4 |
0,61 ± 0,05 |
88,42 ± 6,78 |
1,0 ± 0,09 |
|
|
M ± m |
434,4 ± 0,24 |
0,59 ± 0,01 |
88,96 ± 2,37 |
0,97 ± 0,09 |
|
Использованные в работе ЧПСП, отображающие фоторецепторные свойства листа, целесообразно применять на практике как спектрофотометрические критерии в мониторинге, экологии природных растительных ресурсов и сельскохозяйственных растений.
Библиографическая ссылка
Колдаев В.М. СПЕКТРЫ ПОГЛОЩЕНИЯ ЭКСТРАКТОВ ИЗ ЗЕЛЕНЫХ ЛИСТЬЕВ РАСТЕНИЙ РАЗНЫХ ЖИЗНЕННЫХ ФОРМ // Международный журнал прикладных и фундаментальных исследований. 2018. № 5-1. С. 170-174;URL: https://applied-research.ru/ru/article/view?id=12237 (дата обращения: 19.07.2026).