

Европейский хариус (Thymallus thymallus), как и все лососевые рыбы, обладает ценными вкусовыми качествами, поэтому в последнее время растет интерес к хариусу не только как объекту любительского лова, но и виду, важному для аквакультурного разведения. В некоторых регионах России численность природных популяций европейского хариуса сокращается и этот вид, являясь не только объектом хозяйственного интереса, но и важным звеном водных экосистем, внесен в Красную книгу этих регионов. Рациональное использование, искусственное разведение и охрана вида невозможны без применения современных генетических методов. При этом хариус является объектом не только прикладных, но и фундаментальных исследований. Одним из таких важных направлений являются филогенетические исследования, на базе которых мы можем уточнить положение вида в современной систематике и предположить пути его расселения. Несмотря на то, что в последнее время в научной литературе интерес к этому виду достаточно высок [3–5, 7–9], в российской части ареала большинство популяций остаются неисследованными, данные фрагментарны и затрагивают отдельные реки или их притоки [3, 5, 7, 9].
Соответственно, в задачи нашего исследования входило: уточнить филогенетическое положение европейского хариуса на территории Европейского Севера, а также предположить возможные пути его расселения. Для выполнения этой задачи в работе анализируется контрольный регион мтДНК, относящийся к некодирующим последовательностям мтДНК и относительно быстро накапливающий замены.
Материалы и методы исследования
Всего было проанализировано 60 особей хариуса из разных рек бассейна Белого моря. Географическое расположение выборок показано на рис. 1. Выделение ДНК из плавников проводили с использованием набора QIAGEN DNeasyTM по протоколу производителя. Для амплификации фрагментов использовали последовательности праймеров LRBT–25, LRBT–1195 [8]:. Амплификацию проводили в 15 мкл смеси, содержащей 10ХTaq буфер, 2 мМ Mg2+, 2.5 мM dNTP’s, 1ед. Taq полимеразы (ДИАЛАТ, Россия), 5 пкM каждого праймера, 1.5 мкл ДНК. Амплификацию проводили в термоциклере Tetrad 2 (Bio–Rad Laboratories, Inc., США) в следующем режиме: I – 95 °С 5 мин., затем 35 циклов II – 94 °С 20 сек., 30 сек. 58 °С, 72 °С 1мин., заключительная элонгация 72 °С – 10 мин. Анализ нуклеотидной последовательности проводился в генетическом анализаторе ABI PRISM 3500 (Applied Biosystems, США). Первичная обработка и множественные выравнивание нуклеотидных последовательностей проводили в программе Geneious® 6.0.5, статистический и филогенетический анализы проводили в программах DnaSP V.5 [6], Geneious® 6.0.5 (Biomatters Ltd.). Для построения дендрограммы были использованы последовательности контрольного региона мтДНК, депонированные в базе нуклеотидных последовательностей GenBank® (NCBI).
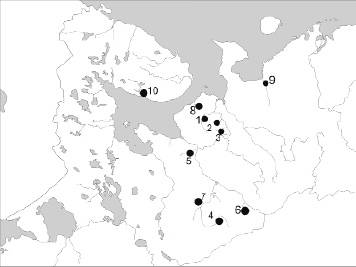
Рис. 1. Карта расположения выборок. Притоки р. Кулой: 1 – Сояна, 2 – Лака, 3 – Келда; притоки р. Северная Двина: 4 – Устья, 5 – Обокша, 6 – Кестваж, 7 – Леменьга; 8 – р. Мегра; 9 – р. Ома; 10 – р. Варзуга
Результаты исследования и их обсуждение
После множественного выравнивания полученных последовательностей исследован участок мтДНК длиной 1046 п.н., включающий полную последовательность контрольного региона Д–петли, GC–содержание этого фрагмента составляло 35.6 %. Было выявлено 13 полиморфных сайтов (1,2 % от всех сайтов), из них информативных 9. Из 14 замен 1 трансверсия, остальные транзиции, также выявлена одна делеция. Среди исследованных последовательностей выделено 10 гаплотипов, 8 – впервые. Распределение гаплотипов по выборкам представлено в таблице.
Распределение гаплотипов в исследованных выборках хариуса
|
Выборки |
Hap1 Main |
Hap2 Ku1 |
Hap3 Ku2 |
Hap4 Ku3 |
Hap5 Meg |
Hap6 ND1 |
Hap7 Ku4 |
Hap8 Ku5 |
Hap9 ND2 |
Hap10 Var |
|
река Кулой |
||||||||||
|
р. Келда |
8 |
2 |
||||||||
|
р. Лака |
14 |
1 |
1 |
1 |
||||||
|
р. Сояна |
11 |
1 |
||||||||
|
река Северная Двина |
||||||||||
|
р. Устья |
1 |
|||||||||
|
р. Обокша |
2 |
|||||||||
|
р. Кестваж |
2 |
6 |
||||||||
|
р. Леменьга |
1 |
1 |
||||||||
|
река Мегра |
2 |
1 |
||||||||
|
река Ома |
1 |
|||||||||
|
река Варзуга |
4 |
Гаплотип, обозначенный нами «Main», выявлен в большинстве выборок за исключением выборки из реки Варзуга. В реке Кулой было обнаружено 6 гаплотипов, 3 из которых являются вариантами основного гаплотипа с однонуклеотидными заменами (рис. 2), а 2 других гаплотипа (Ku4 и 5) с большим количеством замен. Гаплотип Ku5 соответствует гаплотипу AF522430 (At6) (рис. 3), обнаруженному в выборке из бассейна Балтийского моря [9]. В реке Северная Двина было выявлено 3 гаплотипа, гаплотип ND1 относится к группе массового гаплотипа «Main»с одной заменой, а гаплотип ND2 на 5 замен отличается от массового гаплотипа «Main». Гаплотип ND2 (рис. 3) аналогичен гаплотипу AF522426 (At2) выявленному в реке Тана на севере Норвегии [9]. В популяции реки Мегра присутствует гаплотип Meg, отличающийся от основного на 2 замены. В реке Варзуга обнаружен только 1 гаплотип Var, характерный только для рыб из этой реки, отличающийся от массового гаплотипа на 7 замен (рис. 2) и относящийся к той же группе гаплотипов, что и ND2 (рис. 3).
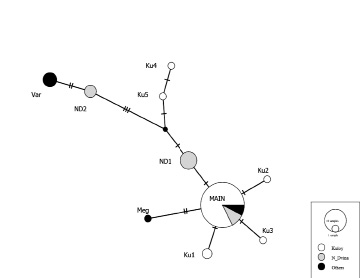
Рис. 2. Сеть гаплотипов контрольного региона мтДНК исследованных рыб на основе алгоритма TCS [2]. Белым цветом выделен Кулой, серым – Северная Двина, черным – выборки из других рек. Обозначения гаплотипов: Ku – Кулой, ND – Северная Двина, MAIN – наиболее распространённый гаплотип, Meg – Мегра, Var – Варзуга
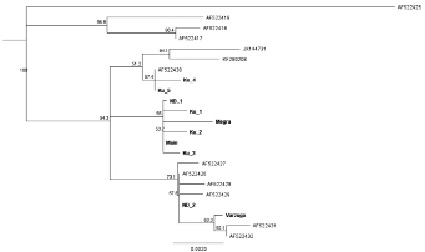
Рис. 3. Филогенетическое дерево гаплотипов контрольного региона мтДНК североатлантической группы европейского хариуса. Дерево построено на основе алгоритма программы Mr Bayes 3.2.1. методом присоединения соседей с использованием HKY модели, в качестве корня использован гаплотип из реки Луары (AF522425). В узлах дерева указаны значения бутстрепа. На дендрограмме гаплотипы, полученные в нашей работе, обозначены Ku_ – Кулой, ND_ – Северная Двина, Main – наиболее массовый гаплотип
На основе проведенного анализа полученные нами гаплотипы относятся к выделенной в предыдущих исследованиях скандинавской филогенетической линии европейского хариуса и образуют три клады, предположительно отражая рефугиумы, из которых происходило расселение хариуса (рис. 3). Во время максимального оледенения северные реки, по мнению Гросвальда [1], сбрасывали воды на юг, соответственно, формирование русел рек Европейского Севера происходило после отступления ледника и расселение хариуса происходило по мере формирования русел. Наиболее вероятно, в период покровного оледенения вид мог сохраняться в сети приледниковых озер, служивших рефугиумами для вида. Большинством исследователей [4, 7] принято допущение, что у холодноводных видов рыб различия в Д–петле достигают 1 % за 1 млн лет. Различия между гаплотипами из разных клад в нашем исследовании достигают от 0.4 % до 0.9 %, при этом клада с гаплотипами из Варзуги (Var) и Северной Двины (ND2) c высокой бутстреп поддержкой (~ 70 %), что предполагает разделение вида и существование вида во время последнего максимального оледенения по крайней мере в двух рефугиумах. Мы можем предположить существование западного рефугиума, откуда происходило заселение хариусом водоемов Фенноскандии и северо-западных притоков Северной Двины. Другим возможным местом рефугиума мог быть расположенный на юго-востоке, возможно, в районе существовавшего в максимальной стадии последнего оледенения Печорского приледникового озера, откуда пошло расселение хариуса в восточные и северо-восточные реки. В пользу этой гипотезы свидетельствует то, что массовый гаплотип обнаружен во всех исследованных популяциях северо-восточных рек и притоков Северной Двины, тогда как в Варзуге и других реках Фенноскандии этот гаплотип и его варианты обнаружены не были. Судя по полученным результатам, предполагаемый рефугиум для популяций Камы и Урала также отличался от этих двух, так как там тоже не был обнаружен гаплотип, наиболее часто встречающийся в наших выборках, а также эти гаплотипы, выявленные в работе Марича с коллегами [7], достаточно сильно (~ 0.5 %) отличаются от остальных гаплотипов скандинавской линии. Для более точного понимания картины расселения европейского хариуса на территории Европейского Севера необходимо более полное исследование выборок не только из рек Арктического побережья, но и притоков верхней Волги.
Заключение
В работе проанализирован фрагмент мтДНК, включающий полную последовательность контрольного региона, у 60 особей европейского хариуса из выборок рек бассейна Белого моря. Было выявлено 10 гаплотипов, 8 из которых были описаны впервые. На основе проведенного анализа все гаплотипы относятся к скандинавской филогенетической линии и образуют три клады, одна из которых с высокой бутстреп поддержкой, что позволяет предположить расселение европейского хариуса на территории Европейского Севера по крайней мере из двух рефугиумов.