

У многих видов покрытосеменных существует апомиксис – формирование семян, в которых зародыш образуется из неоплодотворённой яйцеклетки в результате партеногенеза, а эндосперм из оплодотворённых (псевдогамный апомиксис) или неоплодотворённых (автономный апомиксис) полярных ядер зародышевого мешка. Существует точка зрения, что апомиксис может быть полезен для селекции, так как потомство апомиктов полностью сохраняет удачные комбинации генов в ряду поколений [1, 2]. В то же время есть мнение, что апомиксис полностью отсутствует у диплоидных видов [3], а большинство культивируемых видов являются именно диплоидами.
Одним из таких возделываемых видов является сорго (Sorghum bicolor (L.) Moench), у которого с использованием генетических и эмбриологических методов было показано формирование семян на основе апомиксиса [4–8]. Некоторые из этих работ были позднее подвергнуты критике [3].
Цель данного исследования – выявление нередуцированного апомиксиса у диплоидных линий сорго.
Известно, что у подавляющего большинства видов покрытосеменных растений жизнеспособные семена развиваются при условии, что в эндосперме содержится два набора материнских хромосом и один набор отцовских хромосом (2м:1о). У злаков при нарушении такого соотношения родительских геномов развиваются щуплые зерновки с неразвитым эндоспермом, способные к прорастанию лишь в редких случаях [9]. Однако при опылении нередуцированного зародышевого мешка материнской формы, в котором два диплоидных полярных ядра оплодотворяются диплоидным спермием тетраплоида, возникает гексаплоидный эндосперм со сбалансированным числом хромосом 2м:1о. Поэтому для выявления у диплоидных линий сорго нередуцированных зародышевых мешков, на основе которых могут формироваться апомиктичные семена, нами было предложено в качестве опылителей использовать тетраплоидные линии. В качестве материнских родителей в этом исследовании было решено использовать диплоидные линии, лишенные собственной фертильной пыльцы (линии со стабильной цитоплазматической мужской стерильностью, ЦМС). Для доказательства участия отцовского генома в формировании эндосперма у семян, развившихся на основе псевдогамного апомиксиса, был использован электрофоретический анализ белков эндосперма, как известно, являющихся надежными генетическими маркерами [10, 11].
Материалы и методы исследования
Растительный материал. В качестве материнских родителей использовали ЦМС-линии зернового сорго (Sorghum bicolor (L.) Moench) 9Е Желтозёрное-10 (Ж10) и А3 Ж10, несущие разные типы стерильных цитоплазм: 9Е и А3 соответственно. Проростки этих линий лишены антоциановой окраски. В качестве пыльцевых родителей использовали автотетраплоидные линии АС-1-30-тетра (АСТ) – тетраплоидная линия, полученная на основе линии АС-1, происходящей от регенеранта из культуры тканей, и НТ-3366/2 (НТ) – автотетраплоидная линия, индуцированная колхицином у образца Негритянское кремово-бурое к-3366/2. Проростки этих отцовских линий обладают интенсивной антоциановой окраской. Метелки материнских растений, участвовавших в скрещиваниях, тщательно изолировали до цветения и опыляли по достижении цветения нижнего яруса. Контролем служили изолированные неопылённые метелки. У полученных зерновок отделяли часть эндосперма для электрофоретического анализа спектров запасных белков, а другую часть зерновки с зародышем помещали для прорастания на влажную фильтровальную бумагу. Полученные проростки высаживали в вегетационные сосуды, а после укоренения растения переносили в поле.
Цитологический анализ. Для определения плоидности полученных проростков фиксировали кончики корешков в ацетоалкоголе (1:3) и окрашивали в 4 % ацетогематоксилине после проведении через серию растворов в следующей последовательности: соляная кислота 3,6 % (1N) – 10 мин.; соляная кислота 50 % – 20 мин.; дистиллированная вода – 1–2 мин.; уксусная кислота 45 % – 20 мин; ацетогематоксилин 4 % – 30–60 мин.; дистиллированная вода – 1–2 мин.; смесь мацерирующих ферментов цитаза – 1,5 часа. Временные препараты готовили с использованием смеси 45 % уксусной кислоты и 70 % хлорал-гидрата (1:1), подкрашенной ацетогематоксилином.
Анализ запасных белков эндосперма семян. Белки эндосперма семян анализировали методом SDS-электрофореза в полиакриламидном геле (ПААГ). Острым инструментом отделяли часть эндосперма. Опыт показал, что одинаковые результаты получаются при использовании как сухих, так и замоченных (не более чем на 24 часа) зерновок. Электрофорез в ПААГ проводили в денатурирующих условиях по методу Лэммли. К образцам измельченных эндоспермов добавляли равный объем буфера следующего состава: 0,125 М трис·НСl (рН 6,8), 4 % SDS (додецилсульфат натрия), 20 % глицерин, 0,05 % 2-меркаптоэтанол. Образцы прогревали на кипящей водяной бане (5 мин), после чего вносили в лунки с 4 % концентрирующим гелем. Разделяющий гель содержал 12,5 % раствор акриламида. Электрофорез проводили при постоянном напряжении 70 В. Гели окрашивали раствором нитрата серебра. Для этого гели выдерживали в растворе тиосульфата натрия (0,2 г/л, 1 мин); промывали водой 3 раза по 20 сек. и оставляли на 20 минут в 0,2 %-ном растворе нитрата серебра с добавлением формалина (750 мкл/л). Затем гели промывали водой 2 раза по 20 сек. и проявляли в растворе следующего состава: 6 г карбоната натрия, 50 мкл формалина, 0,4 мг тиосульфата натрия в 100 мл раствора.
Статистическую обработку данных проводили путем сравнения долей по методу Фишера, применяющемуся для малых, а также неравновеликих выборок [12].
Результаты исследования и их обсуждение
ЦМС-линии, использованные в данной работе, отличались стабильностью в течение ряда лет. Однако в некоторых случаях на изолированных неопылённых метёлках отмечали одиночные зерновки. В дальнейшем из них развивались диплоидные мужски-стерильные растения материнского типа.
При опылении метёлок ЦМС-линий пыльцой тетраплоидов число метёлок, на которых завязались семена, статистически значимо возросло; наблюдались метёлки, на которых завязалось от 10 до 70 зерновок, что никогда не наблюдалось в контроле (табл. 1).
Анализ потомства, полученного от опыления диплоидных ЦМС-линий пыльцой тетраплоидов, выявил гибридные и матроклинные (фенотипически полностью сходные с материнским родителем) диплоидные растения (табл. 2). Матроклинные растения были полностью идентичны материнской линии по высоте, окраске листьев, жилки листа, форме метёлки, остистости, окраске колосковых чешуй и пыльников (у ЦМС-линий, включённых в эксперимент, пыльники внешне не отличаются от фертильного аналога, но не растрескиваются при цветении).
Зерновки с гибридным диплоидным зародышем возникают в таких скрещиваниях за счёт гаплоидных пыльцевых зёрен, которые, как это было показано ранее, регулярно образуются у некоторых автотетраплоидных линий сорго, возможно, за счёт соматической редукции [13]. В пыльце тетраплоидов, использованных в нашем эксперименте, содержались пыльцевые зёрна, размер которых соответствует гаплоидным пыльцевым зёрнам, что подтверждает сделанное предположение.
Таблица 1
Результаты опыления ЦМС-линий сорго пыльцой тетраплоидов
|
Комбинация скрещивания |
Число проанализированных метёлок |
Число метёлок с выполненными зерновками |
||||
|
всего |
1–52 |
6–15 |
16–30 |
≥ 30 |
||
|
А3 Ж10, контроль |
23 |
2 ( 8,7 %)1 |
2 |
– |
– |
– |
|
А3 Ж10 × НТ |
19 |
3 (15,8 %) |
3 |
– |
– |
– |
|
А3 Ж10 ×АСТ |
4 |
2 (50,0 %)** |
2 |
– |
– |
– |
|
9Е Ж10, контроль |
49 |
9 (18,4 % ) |
9 |
– |
– |
– |
|
9Е Ж10 × НТ |
17 |
7 (41,2 %)* |
47 |
– |
3 |
1 |
|
9Е Ж10 × АСТ |
8 |
8 (100,0 %)*** |
2 |
2 |
2 |
2 |
Примечание. 1 процентное отношение к числу опылённых метёлок; 2 число зерновок на метёлке; *, **, *** P ≥ 0,95, Р ≥ 0,99, Р ≥ 0,999, по сравнению с контролем (неопылёнными метёлками), в соответствии с F-критерием.
Таблица 2
Характеристика потомства, полученного при диплоидно-тетраплоидных скрещиваниях у сорго
|
Комбинация скрещивания |
Число диплоидных растений |
||
|
всего |
матроклинных |
гибридных |
|
|
9Е Желтозерное 10 × НТ |
51 |
49 (96,1) 1 |
2 |
|
9Е Желтозерное 10 × АС-тетра |
57 |
24 ( 42,1) |
33 |
|
А3 Желтозерное 10 × АС-тетра |
3 |
1 ( 33,3) |
2 |
Примечание. 1 процентное отношение к числу опылённых метёлок.
Матроклинные растения в таких скрещиваниях могли возникнуть а) за счет спонтанного восстановления фертильности в отдельных цветках метёлки; б) за счёт автономного апомиксиса; в) за счет псевдогамии (формирования семени, в котором зародыш образуется путём партеногенеза неоплодотворённой яйцеклетки, а эндосперм – в результате оплодотворения центральной клетки зародышевого мешка). При опылении нередуцированного зародышевого мешка материнской формы, в котором два диплоидных полярных ядра оплодотворяются диплоидным спермием тетраплоида, возникает гексаплоидный эндосперм со сбалансированным числом хромосом 2м:1о.
Анализ генетической природы эндосперма дает возможность уточнить его происхождение. Для этой цели использовали электрофоретический анализ спектров запасных белков, которые у злаков являются четкими генетическими маркерами [10, 11]. Наличие в эндосперме белков, свойственных исключительно материнской форме, свидетельствует об его образовании за счет самоопыления либо автономного развития. Выявление в эндосперме белков, свойственных отцовской форме, или появление новых белков в результате взаимодействия родительских генов в гетерозиготе свидетельствует о гибридной природе эндосперма. В наших исследованиях в электрофоретических спектрах зерновок, завязавшихся в диплоидно-тетраплоидных скрещиваниях, из которых развились матроклинные растения, были выявлены белки, характерные для отцовского родителя, но отсутствовавшие у материнской линии, а также белки, отсутствовавшие у обоих родителей, что указывало на гибридную природу эндосперма этих зерновок и свидетельствовало об их происхождении путем псевдогамного апомиксиса. На рисунке представлены результаты SDS-электрофореза белков эндосперма материнской ЦМС-линии 9Е Желтозерное 10 (1 трек), тетраплоидной линии-опылителя НТ (4 трек) и семян, из которых развились растения материнского типа, завязавшихся на метелке ЦМС-линии 9Е Желтозерное 10, опыленной пыльцой линии НТ (треки 2, 3). Результаты электрофореза наглядно демонстрируют, что в эндосперме зерновки, из которой получено растение материнского типа (трек 2), присутствуют полипептиды, отличающие его от эндосперма материнской ЦМС-линии и указывающие тем самым на его гибридное происхождение. В то же время электрофоретический спектр белков эндосперма другой зерновки, давшей растение материнского типа (трек 3), соответствовал спектру белков материнской ЦМС-линии.
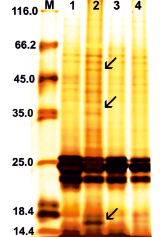
Электрофореграмма запасных белков эндосперма зерновок сорго: 1 – 9Е Желтозёрное-10; 2, 3 – зерновки, завязавшиеся от скрещивания 9Е Желтозёрное-10 х НТ-3366/2; 4 – НТ-3366/2; М – маркеры молекулярной массы. Стрелкой отмечены белки, не свойственные материнской линии
Таким образом, полученные результаты свидетельствуют, что у некоторых зерновок, давших растения материнского типа, эндосперм образовался в результате оплодотворения центральных ядер зародышевых мешков, что свидетельствует о формировании этих зерновок путем псевдогамного апомиксиса. Развитие других зерновок, давших растения материнского типа, у которых электрофоретические спектры белков эндосперма были идентичны материнской линии, могло произойти на основе автономного апомиксиса. Ранее у линии сорго АС-1а при эмбриологическом исследовании выявлены апоспорические зародышевые мешки и автономные зародыши. При опылении этой линии пыльцой диплоидной линии Волжское-4 восковидное, гомозиготной по генам Rs, обусловливающим пурпурную окраску колеоптиля и первого листа проростков, были получены растения материнского типа. Примечательно, что в некоторых скрещиваниях частота таких растений соответствовала частоте цитологически выявленных автономных зародышей или апоспорических зародышевых мешков [8].
Интересно отметить, что ранее предпосылки апомиксиса были выявлены у фертильной линии сорго Желтозёрное 10, стерильные аналоги которой использовали в данных экспериментах. Так, у этой линии выявлена относительно высокая частота полиэмбрионии [14]. При этом у некоторых диплоидно-диплоидных двоен близнецовые проростки были расположены под углом 90 или 1800 по отношению друг у другу, что в литературе трактуется как признак апомиксиса [15]. Результаты данного исследования показывают, что способность к апомиктичному развитию семян сохранилась при переносе генома этой линии на цитоплазмы, индуцирующие мужскую стерильность.
Заключение
Полученные данные свидетельствуют о наличии апомиксиса у диплоидных линий сорго. Использование тетраплоидных опылителей является перспективным подходом для выявления апомиксиса и может быть использовано в работе с разными видами покрытосеменных растений.